先进的疫苗免疫调制剂预防和治疗自身免疫性疾病

自身免疫条件:全球健康危机
流行病学研究表明,环境而不是遗传因素负责当前的自身免疫性传染病。一个主要因素是根除寄生蠕虫在过去40年里由于改善公共卫生保健。这种变化与自身免疫性疾病和突出了生存策略由寄生蠕虫在与宿主的进化。自身免疫疾病,最初仅限于工业化国家,现在蔓延全球的公共卫生改善发展中国家。由于这些疾病的有害影响和经济负担,这种情况可能会破坏这些国家的社会结构。
哺乳动物的机制是为了防止胎儿排斥和副作用许多自身免疫性条件改善怀孕期间,但出生后复发。这种策略也被肿瘤细胞,防止肿瘤炎性反应。在哺乳动物宿主生存,寄生虫的模仿这些机制,偏压宿主的免疫系统抗炎Th2免疫反应逃避破坏而导致在宿主的耐受性。事实上,Th2免疫的特点是抗炎细胞因子和抗体的生产,显然是修复细胞损伤引起的炎性Th1免疫反应。因此,肠虫病提出了用于治疗自身免疫,但由于公众健康危害提出了这是一个不可行的命题。其他的建议方法1是那些基于克隆删除、疫苗、纳米粒子与转基因细胞和细胞疗法。但基因疗法有许多未解决的风险,不管他们的成功,他们的高成本限制他们使用到富裕的国家。目前估计有5000万例自身免疫性在美国,70%的是女性,这一数字在增加,这突显出的大小问题。因此,实际的方法来治疗自身免疫主动免疫治疗或疫苗接种,将重建宽容这些自体抗原(s)容易诱发疾病。
预防和治疗自身免疫性疾病的
自身免疫可能引发的病原体,如果它使神秘的自体抗原或抗原交叉反应的自体抗原。这种情况下生成一个破坏性的t细胞介导免疫对自体抗原(s),促炎Th1 / Th17免疫反应恶化的“抗原决定基蔓延,”响应延伸到其他抗原,加重自身免疫2。因此,治疗和预防自身免疫性疾病需要诱导Th2免疫替代促炎症反应和避免组织和器官损伤。科学家们观察疫苗接种作为一个方法来解决这个问题,但迄今未获成功。这些业绩不佳的原因可能是使用的疫苗配方。虽然都花费了大量的精力疫苗的抗原成分,没有给疫苗佐剂,即负责诱导一个宽容的成分或non-tolerant对抗原的免疫应答3。尽管如此,抑制pro和抗炎免疫系统可以达到我)全球免疫抑制使用DNA或蛋白质合成抑制剂,或基于ii)核对基准点的免疫疗法,那块与生物制剂特异性免疫途径,减少但不能消除免疫抑制4。不幸的是,这些方法增加癌症和感染的易感性。
免疫抑制治疗的另一种选择是诱导抗原特异性抗炎免疫;寄生虫所使用的策略建立宽容。这种方法需要单元疫苗有自体抗原(s) +抗炎佐剂诱导Th2而抑制Th1和Th17豁免,即唯一的抗炎免疫。虽然许多自体抗原刺激自身免疫,缺乏抗炎佐剂疫苗发展的障碍3。这种情况恶化,明矾5和油/水乳剂3在这些疫苗,定期使用佐剂,而认为是纯粹的抗炎,并非如此。事实上,根据不同的疾病,交付模式和其他因素,这些佐剂也可能引起炎性免疫。这或许可以解释为什么有这些佐剂的疫苗治疗自身免疫失败,但成功的传染病,后者通常要求促炎症反应是有效的。
免疫耐受和树突细胞
免疫耐受,无法挂载对抗原免疫反应,不能引起免疫抑制,关闭整个免疫系统6。因此,失败的“自然”为自体抗原免疫耐受,可能引起炎性自身免疫。试图恢复“自然”免疫耐受,使克隆无力和/或删除autoreactive T细胞,迄今为止取得了可怜的临床结果3。因此,另一种是建立一个“诱导”对自体抗原免疫耐受。这里,将取代t细胞介导的自身免疫抗炎免疫;这一过程始于树突细胞(DC)的水平。DCs是启动和控制免疫耐受的抗原呈递细胞,通过调制效应T细胞(苔麸)反应。表型,DCs可以分为不成熟(iDC)、成熟(mDC)和耐受性(tDC)表型7。虽然dc的分化是复杂的,几个关键通路部分阐明。idc的接触外源性抗原和“危险”信号,启动一个分化的过程成为mdc能够刺激T细胞对促炎豁免。因此,mdc显示调节生产MHC, costimulatory分子和促炎细胞因子。在某些情况下,idc分化成tDCs,刺激生产的调节性T细胞亚群)诱导宽容,抑制T细胞免疫,防止自身免疫。
分化成tDCs可以使用我获得)生物制剂如细胞因子,例如il - 10, ii)药理学药剂、雷帕霉素、DCs和iii)基因改造;可能需要复杂的处理的方法。值得注意的是,寄生虫诱发宿主宽容通过切换Th1 / Th2 Th17豁免权,使用直流的凝集素受体DC-SIGN8、9。这种受体,通过绑定甘露糖、岩藻糖,诱发mDC或m * dc的成熟,分别;其中m * DCs Th2-associated直流子集吗9。因此,fucosylated聚糖发挥关键作用,诱导宽容,宽容与胎儿或肿瘤,刘易斯X抗原和LNFPIII相关聚糖结合DC-SIGN。这个绑定允许,通过受体介导内吞作用,抗原的细胞内处理DCs的交付。idc m * dc的分化需要激活的TLR4通过脂质和DC-SIGN并发信号,il - 4的存在10、11。因此,m * DCs将上调抗炎细胞因子的表达,如il - 10,而下调促炎细胞因子,如- 2和IL-17,产生Th2-inducing DCs。幼稚T细胞暴露于Th2-inducing m * DCs + il - 4将促进Th2细胞免疫(图1)。
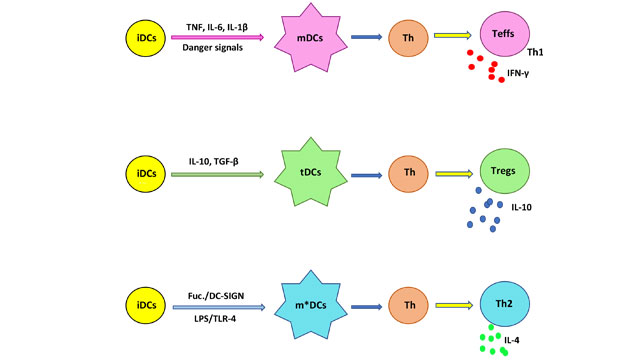
图1所示。成熟的国际数据公司(idc)到不同的子集。暴露在危险的信号,例如TLR配体,和促炎细胞因子,有利于生产的mdc将与辅助T细胞(Th)收益率画眉草,Th1促炎细胞分泌IFN-γ和其他细胞因子。idc的il - 10和TGF-β将成长为tDCs,极化Th细胞亚群,以分泌的抗炎细胞因子il - 10。idc同时刺激通过脂质(有限合伙人)的地和信号的DC-SIGN fucosylated配体(Fuc),将成长为一个m *直流极化Th成为Th2细胞分泌il - 4,这是另一种抗炎细胞因子。
DC-SIGN fucosylated配体,诱导免疫耐受
fucosylated配体在诱导免疫耐受的作用是证明了胎儿和肿瘤宽容;fucosylation过程调节的地方。还提供了额外的证据通过动物研究髓少突细胞糖蛋白(MOG),删除岩藻糖从这糖蛋白导致增加强烈促炎Th17免疫力12。其他例子实验性自身免疫性脑脊髓炎的压制在一个小鼠模型为多发性硬化症(MS)13,在小鼠心脏移植存活时间延长,导致Treg细胞积累14,无论是LNFPIII管理。因此,流行病学和实验证据证实了海藻糖在促进免疫耐受中的作用。作为海藻糖结合DC-SIGN极化对Th2免疫抗原特异性的方式,自身免疫性疾病可能是治疗和预防与自体抗原疫苗和fucosylated聚糖Th2佐剂。同样重要的是,fucosylated聚糖诱导免疫耐受,但没有免疫抑制。这一事实co-delivery dc的抗原和fucosylated聚糖有利于增强亲和力抗体的产生,解释了为什么共价连接fucosylated聚糖诱导出更好的Th2反应抗原。矛盾的是,尽管DC-SIGN8是必不可少的dc的分化和fucosylated的主要受体配体诱导免疫耐受,没有主要的药物开发的努力集中在这个受体作为自身免疫治疗的目标。
为了解决这个需要,新的fucosylated佐剂,qt - 0101,。qt - 0101,炎性辅助QS-21 deacylated导数,是一个定义良好的配糖体的准备,携带岩藻糖残基与刘易斯的无关X抗原,但DC-SIGN能够忍耐15(图2)。qt - 0101引发抗原特异性与抑制Th1 / Th2免疫Th17豁免和亚群的生产15日16,它是与疫苗相关的自身免疫性疾病。因为它的amphiphilicity, qt - 0101可以与蛋白质形成复合物,它允许交付这些抗原的DC被激活和分化向m * DC表型。detergent-like属性将促进早期逃脱的肽蛋白在核内体处理,支持更广泛的一系列构象抗原表位。像fucosylated聚糖,qt - 0101诱导亚群16而抑制促炎豁免权,它不废除它们。因此,与其他经典的疫苗,免疫耐受的维护需要定期免疫刺激免疫系统。暂停接种疫苗,将停止宽容和允许发展的保护性炎性免疫。在需要时恢复炎性免疫的能力,必要的抗感染和癌症,也可以是一个可取的方面的策略,这一过程可以通过重启疫苗接种后情况得到解决。同时,qt - 0101的毒性远远低于其前身QS-21佐剂批准供人类使用。由于成本和方便,疫苗是价值选择生物制剂治疗自身免疫性疾病。然而,他们也可能是有用的防止自身免疫状况,在已知的遗传风险因素存在的情况下。qt - 0101也是一个适合疫苗的佐剂proteopathies,像阿尔茨海默氏症17;需要抗炎的抗体反应,而不是有害的炎性反应。

图2。结构的fucosylated多糖LNFPIII和糖苷qt - 0101。fucosyl残渣(Fuc),蓝色所示,有其羟基3和4 DC-SIGN免费绑定。三萜烯核qt - 0101红色所示。另一个糖基残留黑色所示。
考虑到身体的信息关于qt - 0101的结构活性关系,药物化学的应用方法3、17这种化合物会允许开发新的抗炎药物。在这个领域中做进一步的工作很重要,因为这些化合物的稀缺一直缺乏进展的主要原因为自身免疫性疾病和proteopathies主动免疫疗法。
引用
1。
Carballido JM Santamaria p . 2019。j . Exp。地中海。216:247 - 250
2。
Atassi MZ和Casali p . 2008。自身免疫41:123 - 132
3所示。
Marciani DJ。2017年。自身免疫50:393 - 402
4所示。
李P et al . 2017。前药物杂志。8:460。doi: 10.3389 / fphar.2017.00460
5。
Kooijman年代。et al。J。2018年。蛋白质组学175:144 - 155
6。
青石农协。Immunol。牧师。2011年。241:5-19
7所示。
Maldonado RA和冯Andrian哦。2010年。放置Immunol。108:111 - 165
8。
Geijtenbeek《Gringhuis SI。2009年。启Immunol Nat。9:465 - 479
9。
Hussaarts L。et al . 2014。前面Immunol 5:499。doi: 10.3389 / fimmu.2014.00499
10。
Tundup美国等。2012年。安纽约科学。1253:E1-E13
11。
Gringhuis SI。等。2014年。Nat Commun。5:3898
12。
Garcia-Vallejo JJ。et al . 2014。J Exp地中海。211:1465 - 1483
13。
朱b . et al . 2012。Immunol。142:351 - 361
14。
帕d . et al . 2010。移植90:1071 - 1078
15。
Marciani DJ。2014年。药物。今天19:912 - 920
16。
Heidari对伊朗伊斯兰共和国通讯社表示基于“增大化现实”技术》2019。代数等。乘sci Disord。29:68 - 82
17所示。 Marciani DJ。2019年。研究。DOI: 10.34133 / 2019/5341375

