减少运动神经系统疾病:对神经干细胞特性的影响
 阿达米Raffaella
1
阿达米Raffaella
1
 杰西卡Pagano1
__
杰西卡Pagano1
__
 Michela科伦坡
1
纳塔莉亚Platonova1
黛博拉Recchia2
Michela科伦坡
1
纳塔莉亚Platonova1
黛博拉Recchia2
 Raffaella Chiaramonte
1
Raffaella Chiaramonte
1
 罗伯特·Bottinelli
2
莫妮卡Canepari2
罗伯特·Bottinelli
2
莫妮卡Canepari2
 Daniele Bottai
1
*
Daniele Bottai
1
*
- 1大学卫生科学,米兰,米兰,意大利
- 2帕维亚大学分子医学、意大利帕维亚
宇航员和患者受到慢性movement-limiting病态面对肌肉和/或大脑损伤性能。增加了病人生存的期望和预期的长呆在太空宇航员可能导致长期运动不足和顺向病理的影响。严重的运动限制不仅会影响电机和代谢系统,而且神经系统,改变神经发生和运动和肌肉细胞之间的相互作用。小信息还可以长时间的肌肉停止使用的影响在神经干细胞的特征。我们的在体外研究旨在填补这一空缺通过专注于生物和分子神经干细胞(nsc)的属性。我们的分析表明,nsc来自胡锦涛的SVZ小鼠表现出减少扩散能力和细胞周期的改变。此外,nsc从胡锦涛动物呈现一个不完整的分化和成熟。总体业绩支持的存在之间的联系减少锻炼大脑和肌肉停止使用和新陈代谢,从而代表有价值的新信息,这样既能说明如何情况下如没有负载和缺乏运动,发生在人与一些神经系统疾病,影响nsc的性质和可能导致的负面表现这些条件。
介绍
一些神经系统疾病相关,还是事业,运动障碍;其中,脊髓损伤、多发性硬化、脊髓性肌肉萎缩症与类似的例子对肌肉反重力的影响。同样,众所周知,长时间的太空任务和扩展躺诱导功能改变人体的许多器官,包括骨骼神经肌肉功能的修改Kakurin et al ., 1972)由于反重力肌肉减少活动。
虽然身体活动和认知能力之间的关系已经被几个世纪以来,最近的研究表明自愿体育活动产生的重大影响神经发生(van Praag et al ., 1999;阿达米Bottai, 2016)。自愿体育活动既能产生显著增加增殖祖细胞的水平,恢复神经发生人为的改变在啮齿动物模型的肌肉停止使用/活动(Farioli-Vecchioli et al ., 2014)。自愿的体育活动也已被证明能够克服海马神经发生的年龄相关性损耗(van Praag et al ., 2005)。相比之下,信息相对较少的影响神经发生长时间的肌肉停止使用本身;先前的研究描述在活的有机体内改变很少关注分化过程(Yasuhara et al ., 2007)。因此我们目前缺乏一个详细在体外研究肌肉活动减少的影响在神经干细胞(nsc)特征。
成年神经发生是限于哺乳动物大脑的几个区域:侧脑室的sub-ventricular区(SVZ),它可以检测到评估增殖能力(例如使用标记相关的细胞周期进展如Ki67) (沈et al ., 2008;刘和船员,2017年),sub-granular区海马齿状回的和脊髓(Bottai et al ., 2003)。
外在和内在因素的协同作用在神经源性区域的微环境控制nsc的命运,能够调整未分化的祖细胞之间的平衡和新分化的细胞(Bottai et al ., 2003)。
影响神经发生的决定因素的知识与关键利益的运动限制个人尝试开发新的策略来减少汽车的负面的中部和外围影响固定患者的剥夺和宇航员。长期运动的影响限制在神经发生和营养因素参与的角色这一现象可以使用公认的啮齿动物模型研究的严重运动不足:所谓的后肢卸货(胡)小鼠模型(莫雷et al ., 1979;Desaphy et al ., 2005),再现了突起物上没有重量的支持。在文献中,只有少数研究显示神经生长因子水平的变化(神经生长因子)mRNA和脑源性神经营养因子(BDNF)的躯体感觉皮层,支持假设不用调节神经营养因子表达式(杜邦et al ., 2005)。改变也被证明在大鼠学习能力和记忆受到反重力(太阳et al ., 2009)。胡中央的影响条件包括大幅减少下肢表示的运动皮层鼠(Langlet et al ., 2012)。相比之下,体育锻炼如跑步导致一些祖细胞细胞周期缩短,缩短和s阶段代表的主要内在调节器运行(施加的海马的基础上更上一层的影响Farioli-Vecchioli et al ., 2014)。
低水平的运动被认为是一个主要危险因素的代谢改变(Laaksonen et al ., 2002),可能会影响中枢神经系统,特别是一些神经性的地区(Bottai阿达米、2013;阿达米Bottai, 2016)。L-lactate是一种常见的代谢物在哺乳动物中,其生产发生在所有包括神经元和神经胶质细胞和脑细胞乳酸使用积极的文化(麦地那Tabernero, 2005)。丙酮酸形成在糖酵解和它的一部分转化为L-lactate乳酸脱氢酶(LDH)。这促使我们研究乳酸生产的标记nsc的代谢活动。
我们的研究提供一个新的试验研究,可以补充之前的运动在神经发生的作用。总的来说,我们的分析表明运动在国家安全委员会的角色属性的重要性在体外。
材料和方法
动物模型
实验进行四个月大的雄性小鼠C57BL / 6株。胡诱导电动机剥夺模型我们使用模型(莫雷et al ., 1979;Desaphy et al ., 2005);这表示肌肉停用肌肉萎缩,并适合模仿更改注意到在太空飞行或长期卧床休息。短暂,动物(位于之间的温度20°24°C,湿度35 - 50%和12 h)的光周期暂停单独在笼子里(由实验者模型后Wronski Morey-Holton;Morey-Holton Globus, 2002)通过细绳一端绑到尾巴,另一端连接到笼子的顶部;字符串的长度调整,允许自由移动的动物前肢,虽然身体在30 - 40°倾斜的水平面,这样只前肢触动了笼子的底部。停牌期间(14天),老鼠提供食物和水随意他们的体重每天检查;老鼠的健康状态是确定整个悬架的时期(兽医三次访问的老鼠在卸载实验,第一、7和14天)(见补充材料和方法)。悬挂的第14天,两组的老鼠被牺牲了。
本研究进行了严格按照动物实验伦理委员会的建议在帕维亚大学。动物实验研究符合欧洲法律的实现指令2010/63 /欧盟的欧洲议会和理事会2010年9月22日在动物保护用于科学目的;意大利卫生部授权(授权号码727/2016-PR)的研究项目。所有努力都是尽量减少动物的痛苦;牺牲了在聚氨酯(氨基甲酸)通过腹腔内注射麻醉(1.2克/公斤体重),和老鼠死于颈椎错位。
SVZ的组织学和免疫荧光研究
大脑解剖,冠方前囱和分裂视为中描述Bottai et al。(2014)。简而言之,他们沉浸在4%多聚甲醛在4°C 4 h,冲洗三次,5分钟与磷酸盐缓冲PB(分钟);然后他们被放置在15%蔗糖磷酸缓冲盐PBS 3 h,然后在30% sucrose-PBS过夜。标本被嵌入在最佳切削温度复合(10月),冻结在干冰,切成10μm-thick横向部分通过低温恒温器(徕卡CM1510)。
我们进行免疫荧光染色的研究抗原决定基Ki67 (abcam ab 92353年,1:10 0)的部分与PBS 1 x 15分钟,冲洗孵化NH4Cl 0.05 30分钟,洗了三次与PBS 1 x 5分钟。用于抗原检索样本处理柠檬酸钠10毫米pH值6 5分钟在95°C,在室温下冷却1 h (RT),和洗了三次在PBS 1 x 5分钟。组织被封锁在溶液中含有2%的正常山羊血清(门店),1.25%牛血清白蛋白(BSA)和0.1% Triton X100 90分钟在沿片培养48 h在4°C的溶液0.75%牛血清白蛋白(BSA)和0.05% Triton X100包含Ki67抗体(孵化解决方案)。与初级抗体治疗后,部分与PBS洗1 x和特里同X100 0.05%三次5分钟和15分钟孵化的孵化的解决方案。片被孵化在二级抗体anti-rabbit Alexafluor 488 -共轭(美国免疫学科学IS20012 1:800)解决方案在RT 5 h,复染色和4′,6-Diamidine-2′-phenylindole盐酸盐(DAPI) 300 nM和安装使用FluorSave试剂(Calbiochem)。消极的控制(没有初级抗体)被用来设置为共焦背景水平分析。
数之间的边界是由侧脑室和腹侧的实质SVZ的一部分在300μm从心室最增殖细胞存在面积达0.5毫米2。在前囱层面,两个10μm部分/动物进行了分析,50μm分开,取平均值。阳性细胞的数量之间的选择区域的平均获得了动物组(CTR和胡)用于实验。使用徕卡的图片被收购了TCS SP2共焦显微镜和栈显示为最大强度的预测。在每张图片,我们画一个区域300μm宽,整个长度的SVZ侧壁通过使用徕卡共焦软件2.6版本,我们也计算表面积。
阳性细胞的数量在选择区域获得了动物之间的平均(CTR和胡)用于实验。获得的结果除以平均区域由徕卡共焦测量软件2.6版。研究区域有丰富的丰富的Ki67因为大多数的增殖细胞sub-ventricular区侵入薄壁组织。
大脑的解剖和nsc的准备
中描述的程序执行丹妮拉et al . (2007);Bottai et al。(2008)和Givogri et al。(2008)。简单地说,包含SVZ的大脑被移除和组织被切割出来。每个文化都来自一个鼠标。解剖组织保持在PB 0 01 M方案含有青霉素和链霉素100 U /毫升(表达载体,圣地亚哥,CA)和葡萄糖(0.6%)在4°C的解剖其他样品;然后在37°C执行(酶解离Bottai et al ., 2008)。
组织进行了离心机械解集,直到得到了单个细胞(Bottai et al ., 2008)。最后,上层的丢弃和5毫升的颗粒re-suspended扩散介质(PM) (Gritti et al ., 2002)。在这些条件下,3 - 5天后nsc在组织了球状结构(neurospheres)收获,机械分离和山肩点的浓度10000细胞/厘米2。
扩散分析
增长曲线得到来自六个文化(3 CTR胡和3)从第三段(P3)。在每个通道,细胞机械分离neurospheres何时达到适当的维度(约0.1毫米)。然后他们被机械分离和镀的密度10000细胞/厘米2在一个25厘米2瓶。每个通道的累积的细胞总数计算乘法扩散率(活细胞收获数/培养液细胞数)的累计总数细胞以前的通道(Bottai et al ., 2012)。执行计算的人口倍增时间使用所提供的算法http://www.doubling-time.com。
分化分析
四万细胞被镀成48-multiwell涂板包含一个10毫米(Ricerca Cultrex,特码,意大利)圆形玻璃盖玻片PM中没有EGF 2天,那么这个由点介质中被移除和替换没有EGF、FGF含有1%的胎牛血清。分化后达到7天在37°C公司5%2(Gritti et al ., 2002;Bottai et al ., 2008)。第七天,细胞被洗一次与PBS 1 x和4%多聚甲醛固定(PFA) 10分钟沿主抗体是:使用鼠标anti-β-tubulin三世单克隆抗体(施用,免疫科学ab - 10288);兔子anti-GFAP多克隆抗体(施用、免疫科学ab - 10635)和鼠标anti-O4单克隆抗体(施用、免疫科学马伯- 10259),胞内抗原表位的细胞permeabilized triton X100的0.1%。
二次抗体与荧光团共轭Alexa-fluor 488 (Goat-anti老鼠免疫科学IS20010)和Alexa-fluor 555(山羊anti-rabbit免疫科学IS20012) 1:800的稀释。
共焦分析
用莱卡TCS SP2显微镜图像获得405二极管,他/氖和氩氪激光。
从增殖Neurospheres RNA隔离
为了使用2 * 106细胞RNA制备、750000个细胞被播种在T75厘米血压得到较好的控制2瓶。球体适当的维度时,文化的一小部分机械分离和计数。这一步有可能确定细胞数目大约每毫升的媒介,和文化包含2 * 10的体积6细胞从培养瓶。球体包含介质离心机在100 gs 10分钟和1400年re-suspendedμl无菌PBS 1 x。细胞转移到1.5毫升RNase-free管和离心5000 gs 10分钟。当时颗粒脱落通过地铁和500μl QIAzol裂解试剂(试剂盒)添加到颗粒。细胞混合了1分钟后他们保持在−80°C,直到我们开始准备。
在RT RNA制备样品解冻和RNA净化执行的制造商(见补充材料和方法)。
互补脱氧核糖核酸的合成
互补脱氧核糖核酸制备进行了使用试剂盒(RT2后第一个链工具包)设备,制造商的指示;2μg RNA用于逆转录(见补充材料和方法)。
实时聚合酶链反应
为了研究cell-cycle-regulated基因,使用RT执行实时PCR2分析器(试剂盒)结合RT PCR数组2SYBR绿色Mastermix (pamm - 020佐试剂盒)后MIQE指南(Vandesompele et al ., 2002;泰勒et al ., 2010)。
RT2分析器PCR在96 -孔板阵列包含主要化验84途径集中基因和管家基因(允许规范化的数据),一个基因组DNA控制(检测non-transcribed基因组DNA的污染);3井反转录控制(重要测试逆转录反应)和3井的积极PCR控制的人造DNA序列。中使用的表的基因PCR数组S1报道补充表。
在这个实验中,胡7 CTR和4样品比较(见补充材料和方法)。
CT值归一化所有板分析的基础上获得5最稳定的基因被用作看家基因:到(细胞周期蛋白依赖性激酶4),Cdkn3(细胞周期蛋白依赖性激酶抑制剂3),E2f3 (E2F转录因子3),Itgb1(整合素β1(纤连蛋白受体β)),Shc1 (Src同源性2 domain-containing改变蛋白质C1),板内的5个最稳定的结果。该算法确定这些基因是最稳定的看家基因(补充表S2)。褶皱变化得到2(−ΔΔCT)。数据分析门户网站也产生一个散点图,火山情节,clustergram和热量地图。
流式细胞仪细胞周期分析
Neurospheres来自不同样本(CTR和胡)通道3 - 10机械分离单个细胞,和镀10000细胞/厘米2在两个25厘米2烧瓶为了两届点3和5天的文化和保持在37°C和5%的公司2。
3或5天后,细胞被收获,离心机和resuspended通用缓冲;固定是通过添加乙醇的最终浓度70%,一夜之间(上)在4°C。随后,细胞用PBS 1 x包含的边后卫在5%和离心机在4°C 664 g为4分钟。上层的丢弃,颗粒在PBS resuspended 1 x, 60μg /毫升核糖核酸酶A, 25μg /毫升propidium碘在最后一个25μg /毫升的浓度和0.04% NP40(膜透化作用)。孵化后在4°C在黑暗中,样本获得的FacsVerse流式细胞分析仪和结果进行了分析使用FACSuite软件(BD生物科学)。
代谢研究
MTT试验
细胞被镀上Cultrex(特马Ricerca、意大利)涂层96 -孔板的浓度10000 - 15000细胞/在200点的μl 37°C公司5%2。
一天后电镀和1 h在收藏之前,四唑染色3 - (4 5-dimethylthiazol-2-yl) 2, 5-diphenyl-2H-tetrazolium溴化(MTT)(在PBS 5毫克/毫升;σ)添加到中等(500μg /毫升)的最终浓度。
在37°C, 1 h孵化后点丢弃,通过添加50μl DMSO细胞细胞溶解。在RT 15分钟后,MTT还原测定spectrophotometrically ELISA的读者(日出Tecan)的波长550 nm。作为背景值,我们进行了MTT分析2 h后镀(这一次是附件所需的最小的细胞)。
乳酸生产
使用L-Lactate化验分析设备二世(伊顿生物科学、圣地亚哥、钙、美国)后,制造商的指示(见补充材料和方法)。标准曲线使用L-lactate标准工具包中提供的测量吸光度是执行在570 nm标(日出Tecan)。背景的计算结果都是纠正由所有样本的值减去空白读数。
统计分析
所有数据是正态分布根据Shapiro-Wilk正常测试。所有的数据都表示为代表(AV)±标准错误意味着(SEM)。未配对数据进行评估t以及。结果被认为是显著p < 0.05(见补充材料和方法)。
结果
电动机的Sub-ventricular区剥夺动物显示增殖细胞少于在控制动物
为了评估的影响下肢悬挂SVZ增殖细胞的数量,我们分析了SVZ的胡锦涛和CTR动物比较Ki67的扩散能力测量作为一个表达式如图标志1 a, B。
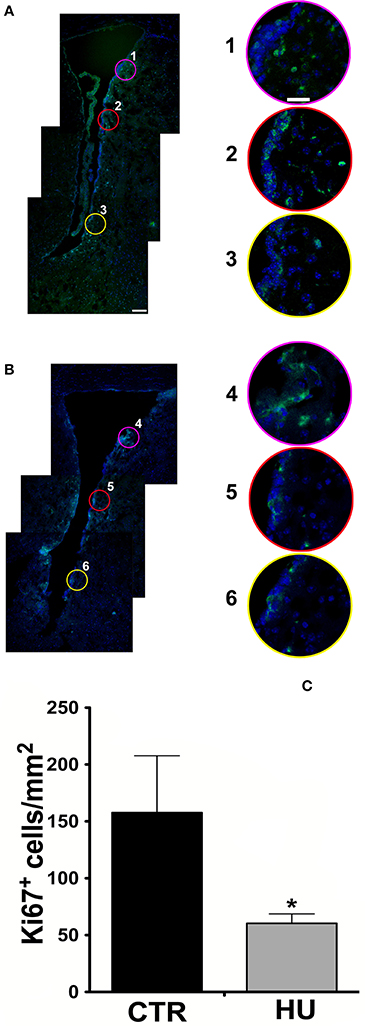
图1。共焦分析10μm日冕部分在前囱级别。Ki67染色(Alexa 488山羊anti-rabbit抗体),用蓝色细胞核与DAPI染色(一)CTR鼠标SVZ(1、2、3是上层的放大,SVZ的中等和较低的地区)(B)胡鼠标SVZ(4、5、6的扩充上,中、低SVZ的地区)。边境的粉色,红色和黄色的放大与圆表示对应较低的放大图(A, B)。(C)每毫米Ki67阳性细胞的数量2在这个地区中描述的材料和方法。价值观代表±SEM手段。我们分析了统计与单侧未配对差异t以及p< 0.0365。象征意义:*p< 0.0365。酒吧100μm规模一个也代表B;比例尺25μm 1代表增大2、3、4、5、6)。
后的工作Yasuhara et al。(2007),为了快速增殖细胞的变化运动约束下,Ki67阳性细胞测定SVZ和腹侧部分的薄壁组织在300μm SVZ的心室,大多数增殖细胞的存在,在一个面积0.5毫米2。在这项研究中,超过3动物被用于组(3 CTR和4胡锦涛在第一个实验中,6 CTR和4胡锦涛在第二个实验中,我们发现了动物细胞增殖减少70%比CTR老鼠(图1 c)。统计分析是由单侧未配对的方式t以及。
歧视的图像被放大的信号,所有的积极Ki67细胞驻留在该地区被计算在内。平均结果分别为157.7±50.0 cell /毫米2CTR和60.4±8.30细胞/毫米2的胡p< 0.0365。CTR的结果(集团)是在协议与那些来自其他作者在老鼠(Azim et al ., 2012),我们推断热图的细胞的数量,和大鼠(Yasuhara et al ., 2007)。
胡小鼠神经干细胞获得显示增殖能力低于CTR老鼠
我们能够隔离nsc两组四个月大的老鼠:那些被停职的尾巴和控制老鼠,它在笼子中自由移动。文化从胡动物出现慢于CTR (neurospheres可比)的形态和增殖的能力在接下来的段落(通过机械分离)显示一个陡坡(图2)。平均斜率,计算曲线的胡锦涛和CTR,明显不同。每个样品的曲线计算;和一个非线性回归(使用Prism项目),获得边坡(起点是抑制250000细胞)。分析表明,平均斜率和标准错误意味着(SEM)分别为0.33063±0.0237N= 3 (CTR平均值±SEM)和0.09753±0.0208N胡= 3。统计分析了使用单侧未配对t以及。的值都明显不同p< 0.0009(图2)。此外,CTR的倍增时间样本(通道(P) 3 - 12)是2.120±0.16天N= 3,而胡锦涛样本(P3-12)倍增时间为7.173±1.434天N= 3,p< 0.0089(图2 b);分析了使用单侧未配对t以及。
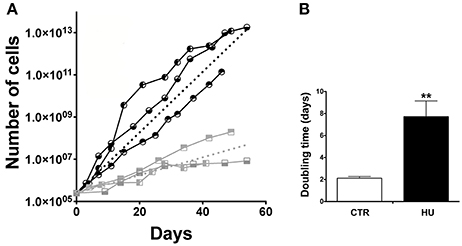
图2。生长曲线和倍增时间。(一)增殖能力比较CTR和胡锦涛样本。曲线的每一个节点代表一个通道。分析了60天的增长。 CTR1;
CTR1; CTR2,
CTR2, CTR3;
CTR3; HU1;
HU1; 胡;
胡; HU3。虚线代表的平均点击率(黑色)和胡锦涛(灰色)。的平均曲线建立的平均点击率曲线的斜率和胡曲线与曲线的约束值250000天在0。(B)倍增时间的扩散中表达天计算的平均倍增时间单一样本通过非线性回归。黑色CTR,灰色。* *p< 0.0089。
HU3。虚线代表的平均点击率(黑色)和胡锦涛(灰色)。的平均曲线建立的平均点击率曲线的斜率和胡曲线与曲线的约束值250000天在0。(B)倍增时间的扩散中表达天计算的平均倍增时间单一样本通过非线性回归。黑色CTR,灰色。* *p< 0.0089。
观察到的差异扩散(类似范围的段落)按照细胞周期的改变。事实上,细胞周期分析、流式细胞分析仪进行显示,细胞数量显著增加在G0 / G1在胡样品(图3):68.95±1.013为胡锦涛和CTR 57.36±1.303,p< 0.0022。与此同时,一个温和的(不是重要的p< 0.1121)减少观察阶段(图3 b在G2 / M)和一个显著降低百分比CTR 20.68±1.212, 13.09±0.9132,p< 0.0075(图3 c)(参见补充图S1)。对于这些实验我们使用4对胡锦涛CTR和样品,实验重复3次,使用双尾未配对分析t以及。
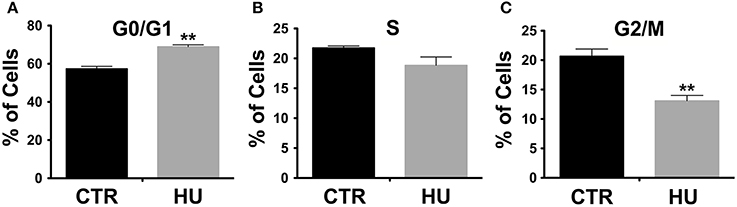
图3。细胞周期分析流式细胞分析仪。显著增加细胞数量存在于G0 / G1在胡* *样品p< 0.0022(一);与此同时轻微下降阶段(B)并在G2 / M比例显著降低(C)是观察到的。* *p< 0.0075(C)。黑色CTR,灰色。
未发现显著差异水平的细胞凋亡与衰老的胡锦涛和CTR nsc(见补充数据)。另一方面,细胞的数量表达巢蛋白,Ki67, GFAP显著改变在胡派生nsc(见补充图S3和补充的结果)。
胡小鼠神经干细胞获得显示分化和成熟能力低于CTR老鼠
我们观察到分化的nsc从暂停获得小鼠受损,显示β-tubulin三世阳性细胞数量显著低于在CTR nsc co-expression胶质原纤维酸性蛋白(GFAP)(数据4 a e),这表明一个不完整的分化和/或成熟的nsc,最有可能的是,不达到成熟神经元膜电气性能。分化神经元的百分比为6.8±1.74(平均值±SEM)NCTR = 4和0.56±0.2N= 4p< 0.0119的胡锦涛样本。同样,胡锦涛的文化产生少突胶质细胞(数据显示较低的能力4 f-h)。差异化的少突胶质细胞的百分比也放弃了在这种情况下,从1.74±0.54N= 3的CTR 0.00±0.01N= 3p< 0.0322的胡锦涛样本。变化的统计分析都使用了双尾未配对的分化细胞t以及。
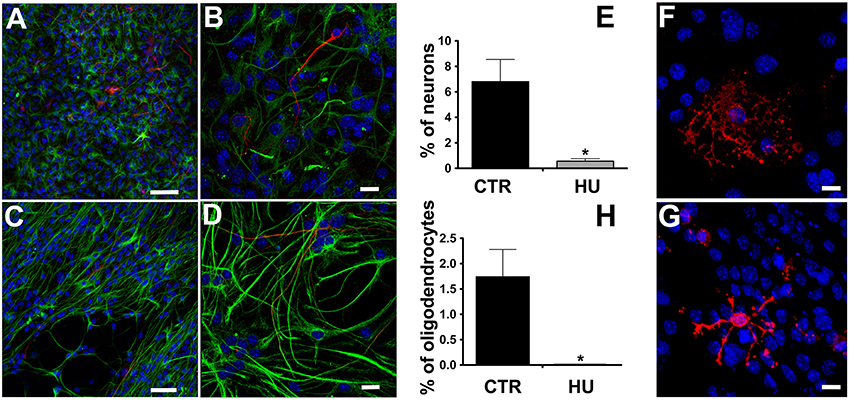
图4。分化细胞组织化学染色。β-tubulin三世(555年红,Alexa山羊anti-mouse抗体)和GFAP(绿色,Alexa 488山羊anti-rabbit抗体),细胞核蓝色,DAPI染色。(一个,C)50μm酒吧。(B, D)更高的放大倍数,20μm酒吧。(E)定量分析β-tubulin三世CTR标记表达式(n= 4)和胡(n= 4)*p< 0.05。O4(红色,Alexa 555山羊anti-mouse抗体),细胞核蓝色,DAPI染色(F)CTR,(G)胡(H)定量分析的CTR O4标记表达式(n= 4)和胡(n= 4)*p< 0.05。20μm酒吧。
暂停动物显示,细胞周期基因表达改变
两个基因被发现是CTR和胡样本之间明显不同:Cdk5监管subunit-associated蛋白1 (Cdk5rap1)和一个折叠−3.53的规定p< 0.005624和细胞周期蛋白依赖性激酶6 (Cdk6)规定2.38折起来p< 0.021001图5(补充图S2和S3补充表)。另一个基因,细胞周期蛋白依赖性激酶抑制剂2 (Cdkn2a),显示褶皱变化调节超过2但这差异不显著(补充图S2和S3补充表)。
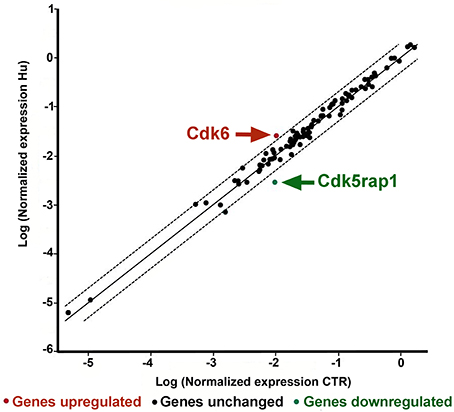
图5。PCR数组CTR和胡样本分析。散点图分析的规范化表达基因在两组之间的数组(CTR和胡)通过密谋反对另一个,说明大的基因表达改变。中央线表明改变基因表达而虚线显示所选的折叠调节阈值。数据点超出虚线在左上和右下部分超过所选折监管门槛。观察到的两个基因之间明显不同CTR和胡锦涛用箭头表示。(控制= CTR,n胡和后肢卸载= = 7日,n= 4)。
胡锦涛和CTR之间其他基因显示一个改变表达式但这些差异低于2倍或不显著。
统计分析是通过双尾未配对的方式t以及。
在Movement-Restrained小鼠代谢改变
众所周知,nsc主要glycolysis-based代谢,而转移到氧化代谢在分化。为了评估和比较两组的生存能力和代谢nsc (CTR和胡),我们执行一个MTT测定和分析产生的乳酸水平的文化。
MTT试验,为了获得代谢活动的起点,我们第一次测量后MTT的水平细胞粘附在威尔斯(之前涂上Cultrex特马Ricerca,意大利)。1或3天后,实验和测量结果减去执行从背景中(以无细胞)和扩展的函数在第0天获得的基线。细胞的细胞生存能力胡(表示为任意单位)明显低于CTR的(后者的前40%),(p在一天的文化(< 0.0364)6)和52% (p< 0.0017)(图3天的文化6 b)。实验进行了三次使用至少5对胡锦涛的CTR和4样品样本。
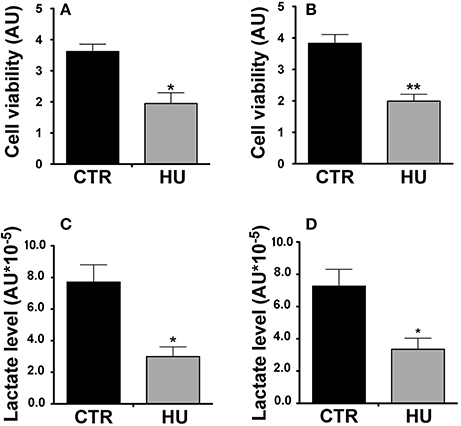
图6。胡锦涛和CTR nsc的代谢活动。MTT试验。(一)细胞生存能力表达任意一天后单元(AU)在体外文化*p< 0.0364。(B)细胞生存能力表现在非盟三天后在体外文化* *p< 0.0017。黑色= CTR;灰色=。(C)乳酸水平:相对乳酸生产中表达非盟* 10−54 h (在体外文化*p< 0.0142。(D)相对乳酸生产中表达非盟* 10−5,18 h (在体外文化*p< 0.0275。黑色= CTR;灰色=。
MTT测定使用双尾未配对进行统计分析t以及。
我们的结果证实了胡样本的代谢能力低;4 h后确实培养胡锦涛样本显示,61.2%乳酸低于CTR样本(p< 0.0142)(图6摄氏度胡)而18 h后培养的样品是CTR的46.1%的水平,p< 0.0275(图6 d)。统计分析是通过双尾未配对的方式t以及。
讨论
严重减少运动存在于许多不同的疾病如脊髓损伤、脊髓肌肉萎缩,肌萎缩性脊髓侧索硬化症,多发性硬化症,可以是由于损伤骨骼肌肉的装置或中枢和周围神经系统。这个条件也是常见的床靠背病人(由于各种病态)或宇航员经历长时间的低重力。减少运动或重力刺激产生对人体的重要作用,改变许多器官,包括大脑的活动。研究这些影响,我们利用下肢卸载协议(莫雷et al ., 1979;Desaphy et al ., 2005),它再现了减少运动或在太空失重。
我们的经验与胡锦涛的老鼠,用于现在和其他的研究(Brocca et al ., 2010;Desaphy et al ., 2010;马费伊et al ., 2014;Cannavino et al ., 2015),表明胡可以产生巨大的压力在大约6%的情况下,在第一次暂停3天(【et al ., 1994;Morey-Holton Globus, 2002;正在et al ., 2010;Gaignier et al ., 2014;Mysoet et al ., 2014),之后的水平压力,血清,尿液测量皮质甾酮,回到正常的non-suspended动物。事实上,它已经证明(【et al ., 1994),在5-week-old对照组大鼠皮质甾酮尿排泄是87.4±12.8 ng /天,并没有改变整个卸货时间。显著增加(超过三倍)在发现皮质甾酮排泄天1和3在暂停之前回到控制水平,这次小长在老年大鼠(【et al ., 1994)。与此同时,增殖细胞的数量在鼠神经源性领域不受肾上腺切除术,治疗这些动物糖皮质激素诱导海马的显著增加,但不是SVZ (阿隆索,2000)。其他作者(李et al ., 2016)表明,慢性肾上腺酮的注射,28天,可以显著降低SVZ的细胞增殖和分化。然而,这种治疗模仿很长应力状态这不是类似于我们的模型。事实上,即使在21天之后,胡锦涛老鼠没有任何明显的皮质甾酮不同等离子体水平对控制老鼠(Gaignier et al ., 2014)。
胸腺和脾脏重量由体重(归一化)和脾脏的有核细胞的数量证明没有统计不同CRT和胡(Gaignier et al ., 2014)。在我们的经验中,我们没有发现差异的重量肾上腺(体重标准化)暂停和CTR老鼠(Bottai、Recchia Canepari未发表的结果)。
这些数据表明,暂停所有动物,可以自由探索笼子里没有攀爬,接受小压力和最有可能nsc的变化特点是由于减少了整体的运动,下肢肌肉的选择性停止使用。尽管胡锦涛回老鼠四肢是免费自愿通过全方位的运动运动,四肢肌肉不暴露在重负荷和缺乏一个活跃的肌肉力量和力量的发展。
许多假说可以解释胡nsc性质的作用。后肢骨骼运动的减少可能引起的变更从肌内感受器传入信号和反馈信息,大脑皮层,由于反射组织的修改在下肢肌肉群(D 'Amelio et al ., 1998)。同样,长期单侧下肢悬挂,在人类中,引起神经系统的一种变体塑料属性(克拉克et al ., 2006)。在网站上的神经递质,它是证明28天胡暴露大鼠诱导和空间学习记忆功能的变化相关蛋白表达的变化(75蛋白过度表达和72年衰减;王et al ., 2017)。特别是,谷氨酸受体的表达(GluR) 1, GluR4,谷氨酸水平的差异(王et al ., 2017)。另一方面,5 -羟色胺的浓度,多巴胺,GABA,肾上腺素是减少对控制大鼠(王et al ., 2017)。其他研究表明,HU-induced mRNA水平改变营养的因素如神经生长因子、脑源性神经营养因子的躯体感觉皮层(杜邦et al ., 2005)。此外,许多因素,如细胞因子和多肽,而且代谢物,液,和microrna在循环系统发布的肌肉活动期间允许可能影响其他组织(即。、骨骼、肝脏、脂肪组织、心血管系统,大脑)与自分泌、旁分泌和内分泌的影响(2011年彼得;Giudice和泰勒,2017年;Pourteymour et al ., 2017;Safdar和线粒体,2018;Whitham et al ., 2018)。最后,其他作者曾表明,抑制骨形成,由卸荷,是下肢海拔的结果独立于糖皮质激素之间的血浆皮质酮水平没有显著的不同通常加载和卸载老鼠和肾上腺切除术没有保护作用在卸载骨质疏松动物(Halloran et al ., 1988)。
在我们之前的经验,老鼠拒绝吃前两天暂停他们解开第二天和丢弃的研究。在当前的研究中,没有一个动物被丢弃。我们以前的观测表明,动物的体重进行悬架有轻微但体重都明显下降(Desaphy et al ., 2010),而其他研究人员即使长时间悬挂相比对照组(没有发现差异Colaianni et al ., 2017)。
在卸载实验,小鼠为适当的食物和水的摄入,每天监测排尿和排便正常的梳理行为,观察是统计两组之间的相似,表明他们经历很少的压力。权重的无重量的肌肉(腓肠肌和比目鱼肌)显著降低(分别为10和20%)卸货后,作为我们集团已经在以前的工作证明(Brocca et al ., 2010从其他组()和汉森et al ., 2013)。
我们的研究增加了更多的信息,更好地理解运动的作用减少nsc特性。众所周知,缺乏身体活动是一个风险因素对阿尔茨海默病(AD) (桥本et al ., 2017)由于海马萎缩相关广告。Guure和合作者在最近的荟萃分析发现,体育锻炼预防广告比所有其他形式的痴呆(Guure et al ., 2017)。
体力活动引起的海马体积的增加和改善神经发生(Bednarczyk et al ., 2009)最有可能通过增加血流量(卡斯,2017)。一致,暂停大鼠的血浆VEGF水平下降后14天的治疗,而在大脑中仍然是不变的(Yasuhara et al ., 2007)和比目鱼肌(Wagatsuma 2008)。长暂停(4周)诱导炎症反应在大鼠颈总动脉暴露于模拟微重力,表明炎症反应可能是细胞动脉重塑机制,负责在暴露于模拟微重力(刘et al ., 2014)。前瞻性研究表明,缺乏身体活动是最常见的可避免的风险因素之一,发展广告。此外,身体活动水平升高与降低广告的风险相关联。广告病人进行了运动训练减少神经精神症状,改善认知功能和日常生活的活动缓慢下降(卡斯,2017)。运动后适当广告后病人恢复了一些的大脑功能和控制体能训练(细语et al ., 2014)。
我们决定研究SVZ神经发生在成年人因为它的作用might-albeit没有可比性,在活动期限,rodents-have对人类大脑的健康产生重大影响。现在已经知道,成神经细胞来源于SVZ通过吻侧迁移流系统迁移到嗅球(Sanai et al ., 2004;van den Berge et al ., 2010)。然而,有另一个主要的未成熟神经元的迁移途径运往婴儿的前额叶皮层(Sanai et al ., 2011),它被发现,通过测量核弹test-derived的水平14在基因组DNA C (伯格曼et al ., 2012),这是微不足道的人类嗅球神经发生。令人惊讶的是它是确定有产后细胞营业额在成年人的纹状体(恩斯特et al ., 2014),作者建议来源于SVZ。尽管这些证据,有有限的线索锻炼在SVZ神经发生的作用存在争议影响(布朗et al ., 2003;Bednarczyk et al ., 2009;布莱克摩尔et al ., 2012;崔et al ., 2014)。相反,运动的积极作用,特别是在前大脑损伤的情况下,保证(李et al ., 2016;Mastrorilli et al ., 2017)。最后,对海马的影响更广泛,建立了(李et al ., 2016;Mastrorilli et al ., 2017;斯奈德et al ., 2017;Bouchard-Cannon et al ., 2018;弗斯et al ., 2018;Masrour et al ., 2018)。
这促使我们进行一个范围的证据体外分析证明,已经显示在大鼠海马模型(Yasuhara et al ., 2007),下肢卸货产生增殖Ki67的减少(内源性细胞增殖的标志,在细胞核表达细胞周期的所有阶段,除了休息阶段,相关联,且必要的,细胞增殖)阳性细胞在SVZ(图1)。
这第一步允许我们预测一些改变在nsc属性SVZ的暂停动物;然后我们继续准备NSC来自这个地区的文化。
我们能够生产NSC文化SVZ的胡锦涛和CTR老鼠;我们发现胡锦涛nsc neurospheres更很难获得,因为他们出现更慢比控制。neurospheres两个文化群体的形态相似,在形状或粘连的病例中没有区别的塑胶瓶。然而,我们注意到之前做任何计算的大小neurospheres显然是较小的胡组。扩散分析测量细胞数量为许多段落在指数增长阶段。这个过程表明,扩散能力的胡派生SVZ nsc受损,确认的结果体外分析(图2)。我们分析了抗原表位的表达水平通常在nsc发现如巢蛋白、GFAP SOX2 (库恩et al ., 2016),GLAST (Gubert et al ., 2009),Ki67,表示在增殖细胞在细胞周期的所有活动阶段。细胞的数量表达巢蛋白、Ki67和GFAP在胡派生nsc显著降低。减少数量的Ki67阳性细胞nsc增殖(图上支持我们的结果2)。巢蛋白和GFAP阳性细胞减少(补充图S3)可能与更少的犯神经胶质细胞,导致更少的胶质分化也显示在图4。相反,Sox2,这是一个更加值得信赖的干细胞标记(Sarkar Hochedlinger, 2013),没有任何变化的表达式。这个结果可以解释考虑到我们评估阳性细胞的数量,这是与细胞周期动力学的变化,我们观察(图3),而不是减少这些标记每个细胞的表达水平。进一步分析将是必要的,以了解这些方面。
暂停导致逮捕的nsc G0 / G1期,并显著减少G2 / M期(图3),G0 / G1逮捕也发现老鼠bmsc当暴露于模拟或真实微重力(憎恨et al .,以及2014年)。
的影响下肢卸载stereological参数的雄性老鼠的海马和皮层运动区最近评估(见补充讨论)。
另一个有趣的方面是潜在的可逆性变化引起成人SVZ nsc,后或多或少长期post-HU复苏。经济复苏(暂停之后)有或没有锻炼的老鼠无效检索SVZ的增殖和分化(Yasuhara et al ., 2007)。
对胡锦涛和CTR nsc的代谢能力,认识到细胞内的订单2值的0.5托或少发生在O2召集氧化磷酸化和乳酸的生产和积累(Rogatzki et al ., 2015)。在低水平的氧,厌氧代谢提供的能量,导致乳酸的生产(De菲利皮主持和迪莉娅,2011年)。肝星状细胞,干细胞,如ESCs msc、和其他人,不成熟的线粒体形态,减少氧化能力和增强糖酵解途径合成代谢水平(Folmes et al ., 2012)。大脑乳酸氧化代谢的主要底物在开发过程中,选择性地利用水库作为合成细胞增殖和分化。此外,大脑细胞乳酸使用积极的文化(麦地那Tabernero, 2005)。在这个场景中,从我们的工作结果强调的参与代谢改变在运动活动的减少和nsc属性。
MTT试验已被广泛用于评估细胞生存能力;然而,MTT有效措施活细胞的代谢活动,因为它对细胞的能力,减少线粒体功能的四唑染色。氧气的水平已经被证明能够影响nsc特性在正常的人体发育进程中,疾病和培养(瑞士思德利公司et al ., 2000;Mohyeldin et al ., 2010;Santilli et al ., 2010)。自从nsc大多糖酵解代谢,由于低氧张力水平(Zhang et al ., 2015)降低乳酸水平以胡锦涛nsc表明这些细胞氧化代谢(图更高6 c, D),这也证明了其他作者(麦地那Tabernero, 2005)。
与预期的相反,一个基因,Cdk5监管subunit-associated蛋白1 (CDKrap1),显著降低在胡锦涛样本相对于CTR(图5)。Cdk5rap1首次发现负面Cdk5监管机构,但在随后的研究中,发现Cdk5rap1负责post-synthetic RNA修改N6-isopentenyladenosine(我的转换6一)2-methylthio-N6-isopentenyladenosine (ms2我6)。(Cdk5rap1)负责2-methylthio修改的哺乳动物线粒体(mt)转运rna Ser (UCN)法酪氨酸,Trp基码(王et al ., 2015;魏et al ., 2015)。在乳腺癌细胞MCF-7 Cdk5rap1缺乏诱导细胞周期阻滞(王et al ., 2015)。Cdk5rap1-KO老鼠对应激敏感线粒体重建(见补充讨论b)。
另一个基因表达改变在胡锦涛nsc,相比于CTR nsc Cdk6。
我们的研究结果的基础上,我们可以推测,NSC的低表达水平的Cdk6 CTR动物与他们更高的增殖能力,所以这种基因的表达水平的升高在胡派生NSC应该与他们的改变(图的功能5)。Cdk6,作为细胞周期激酶,促进发展S期,染色质状态,细胞死亡,细胞生存,和DNA修复与细胞周期蛋白D交互;此外,行为作为转录监管机构、Cdk6与其他蛋白质相互作用如RUNX1、NF-kB, STAT3,和AP-1分别调节分化、炎症、细胞周期阻滞、压力造血和血管生成(Tigan et al ., 2016)。Cdk6和细胞周期蛋白D是肿瘤发生的非常重要的动力。Cdk6可以作为肿瘤抑制减少增殖淋巴恶性肿瘤(Kollmann et al ., 2013),或者抑制乳腺癌细胞的增殖(卢卡斯et al ., 2004)。进一步分析将必要的描述这方面。
这些数据,据我们所知,第一个证据nsc属性之间的相关性变化的运动约束后,新陈代谢修改和基因表达的变化。有趣,胡锦涛对维持细胞改变属性的能力超过10通道文化表明表观遗传修饰可能参与其中。在这些方面,暂停动物进行了应力卸载的头三天期间可以解释基因表达改变的表观遗传变化。然而,我们进行细胞周期基因基因表达分析显示没有变化影响的基因通常在压力感应即细胞周期蛋白D1和细胞周期蛋白依赖性激酶抑制剂1 (P21) (Juszczak Stankiewicz, 2018),而是其他基因改变。
表观遗传修饰不仅影响DNA和蛋白质,而且编码和非编码rna。rna在超过100种方法包括可以修改N6-methyladenosine (m6A),N7-methylguanosine (m7G) m5C、假尿苷和queuosine (贾et al ., 2013)。许多这样的修改有一个基本的RNA代谢的功能在控制方面,如拼接、交通、翻译、和退化。这些活性RNA修改代表另一个水平的基因调控,称之为“epitranscriptomics。“发现Cdk5rap1(充当methylthiotransferase线粒体RNA)在其表达改变电动机剥夺后是非常有趣的在这种情况下,并打开一个新的神经发生代谢和细胞之间的联系的监管。
作者的贡献
RA和DB:设计研究;RA,摩根大通,MCo, NP,博士,MCa,和DB:进行研究;RA, MCo, NP、RC和DB:分析和解释数据;RA和DB:写论文;RA,摩根大通,MCo, NP,博士,RC, RB, MCa,和DB:修订和批准。
利益冲突声明
作者声明,这项研究是在没有进行任何商业或财务关系可能被视为一个潜在的利益冲突。
确认
稿件的编辑是由Damian反的校对和编辑服务为学术界和业务在英国和欧洲(http://englishedituk.co.uk)。
这项工作是支持的资助从Asamsi DB ONLUS通过Prosciutta 23 - 48018法(RA)、意大利和垂直基金会,通过卡洛Bernari 13日00139罗马(IT),意大利。MCo支持基金会Italiana / la Ricerca南Cancro(18013年博士后奖学金)。
我要感谢教授Riccardo Ghidoni对他的支持。我给罗伯特巴尔迪尼特别感谢。
补充材料
本文的补充材料在网上可以找到:https://www.frontiersin.org/articles/10.3389/fnins.2018.00336/full补充材料
引用
阿达米,R。,和Bottai D。(2016). Movement impairment: focus on the brain.j . >。Res。94年,310 - 317。doi: 10.1002 / jnr.23711
阿隆索,g (2000)。延长皮质甾酮治疗的成年老鼠抑制少突细胞祖细胞的增殖在灰质和白质的大脑区域。神经胶质31日,219 - 231。doi: 10.1002 / 1098 - 1136 (200009) 31:3 < 219:: AID-GLIA30 > 3.0.CO; 2 r
Azim, K。,Fiorelli, R., Zweifel, S., Hurtado-Chong, A., Yoshikawa, K., Slomianka, L., et al. (2012). 3-dimensional examination of the adult mouse subventricular zone reveals lineage-specific microdomains.《公共科学图书馆•综合》7:e49087。doi: 10.1371 / journal.pone.0049087
正在,D。,N我cchia, G. P., Desaphy, J. F., Camerino, D. C., Frigeri, A., and Svelto, M. (2010). Analysis by two-dimensional Blue Native/SDS-PAGE of membrane protein alterations in rat soleus muscle after hindlimb unloading.欧元。j:。杂志。110年,1215 - 1224。doi: 10.1007 / s00421 - 010 - 1592 - 6
Bednarczyk, m R。Aumont,。、Decary年代。,Bergeron, R., and Fernandes, K. J. (2009). Prolonged voluntary wheel-running stimulates neural precursors in the hippocampus and forebrain of adult CD1 mice.海马体19日,913 - 927。doi: 10.1002 / hipo.20621
伯格曼,O。Liebl, J。,Bernard, S., Alkass, K., Yeung, M. S., Steier, P., et al. (2012). The age of olfactory bulb neurons in humans.神经元74年,634 - 639。doi: 10.1016 / j.neuron.2012.03.030
布莱克摩尔,d G。Vukovic, J。,Waters, M. J., and Bartlett, P. F. (2012). GH mediates exercise-dependent activation of SVZ neural precursor cells in aged mice.《公共科学图书馆•综合》7:e49912。doi: 10.1371 / journal.pone.0049912
Bottai D。,和阿达米,R。(2013). Spinal muscular atrophy: new findings for an old pathology.大脑病理学研究。23日,613 - 622。doi: 10.1111 / bpa.12071
Bottai D。,Cigognini, D., Nicora, E., Moro, M., Grimoldi, M. G., Adami, R., et al. (2012). Third trimester amniotic fluid cells with the capacity to develop neural phenotypes and with heterogeneity among sub-populations.备份文件夹。神经。>。30,55 - 68。doi: 10.3233 / rnn - 2011 - 0620
Bottai D。,Fiocco, R., Gelain, F., Defilippis, L., Galli, R., Gritti, A., et al. (2003). Neural stem cells in the adult nervous system.j . Hematother。干细胞Res。12日,655 - 670。doi: 10.1089 / 1525816036073268
Bottai D。,Madaschi, L., Di Giulio, A. M., and Gorio, A. (2008). Viability-dependent promoting action of adult neural precursors in spinal cord injury.摩尔。地中海。14日,634 - 644。2008 - 00077. - bottai doi: 10.2119 /
Bottai D。,Scesa, G., Cigognini, D., Adami, R., Nicora, E., Abrignani, S., et al. (2014). Third trimester NG2-positive amniotic fluid cells are effective in improving repair in spinal cord injury.经验,神经。254年,121 - 133。doi: 10.1016 / j.expneurol.2014.01.015
Bouchard-Cannon, P。洛登,C。陈,D。,和Cheng, H. M. (2018). Dexras1 is a homeostatic regulator of exercise-dependent proliferation and cell survival in the hippocampal neurogenic niche.科学。代表。8:5294。doi: 10.1038 / s41598 - 018 - 23673 - z
憎恨,以及。Barenghi, L。,和Maier, J. A. (2014). Stem cells toward the future: the space challenge.生活4,267 - 280。doi: 10.3390 / life4020267
Brocca, L。,Pellegrino, M. A., Desaphy, J. F., Pierno, S., Camerino, D. C., and Bottinelli, R. (2010). Is oxidative stress a cause or consequence of disuse muscle atrophy in mice? A proteomic approach in hindlimb-unloaded mice.Exp。杂志。95年,331 - 350。doi: 10.1113 / expphysiol.2009.050245
布朗,J。,Cooper-Kuhn, C. M., Kempermann, G., Van Praag, H., Winkler, J., Gage, F. H., et al. (2003). Enriched environment and physical activity stimulate hippocampal but not olfactory bulb neurogenesis.欧元。j . >。17日,2042 - 2046。doi: 10.1046 / j.1460-9568.2003.02647.x
Cannavino, J。Brocca, L。Sandri, M。葛,B。,Bottinelli, R., and Pellegrino, M. A. (2015). The role of alterations in mitochondrial dynamics and PGC-1alpha over-expression in fast muscle atrophy following hindlimb unloading.j .杂志。593年,1981 - 1995。doi: 10.1113 / jphysiol.2014.286740
崔,c . H。,Jung, S. L., An, S. H., Park, B. Y., Kim, T. W., Wang, S. W., et al. (2014). Swimming exercise stimulates neuro-genesis in the subventricular zone via increase in synapsin I and nerve growth factor levels.医学杂志。体育运动31日,309 - 314。doi: 10.5604/20831862.1132130
克拉克,b . C。,Manini, T. M., Bolanowski, S. J., and Ploutz-Snyder, L. L. (2006). Adaptations in human neuromuscular function following prolonged unweighting: II. Neurological properties and motor imagery efficacy.j:。杂志。101年,264 - 272。doi: 10.1152 / japplphysiol.01404.2005
Colaianni G。,Mongelli, T., Cuscito, C., Pignataro, P., Lippo, L., Spiro, G., et al. (2017). Irisin prevents and restores bone loss and muscle atrophy in hind-limb suspended mice.科学。代表。7:2811。doi: 10.1038 / s41598 - 017 - 02557 - 8
D 'Amelio, F。福克斯,r。吴,l . C。,Daunton, N. G., and Corcoran, M. L. (1998). Effects of microgravity on muscle and cerebral cortex: a suggested interaction.Res放置空间。22日,235 - 244。doi: 10.1016 / s0273 - 1177 (98) 80015 - x
丹妮拉,F。,Vescovi, A. L., and Bottai, D. (2007). The stem cells as a potential treatment for neurodegeneration.摩尔。生物方法。399年,199 - 213。doi: 10.1007 / 978 - 1 - 59745 - 504 - 6 - _14
菲利皮主持,L。,和Delia, D. (2011). Hypoxia in the regulation of neural stem cells.细胞。摩尔。生命科学。68年,2831 - 2844。doi: 10.1007 / s00018 - 011 - 0723 - 5
Desaphy, j·F。、Pierno年代。,Liantonio, A., De Luca, A., Didonna, M. P., Frigeri, A., et al. (2005). Recovery of the soleus muscle after short- and long-term disuse induced by hindlimb unloading: effects on the electrical properties and myosin heavy chain profile.一般人。说。18日,356 - 365。doi: 10.1016 / j.nbd.2004.09.016
Desaphy, j·F。、Pierno年代。,Liantonio, A., Giannuzzi, V., Digennaro, C., Dinardo, M. M., et al. (2010). Antioxidant treatment of hindlimb-unloaded mouse counteracts fiber type transition but not atrophy of disused muscles.杂志。Res。61年,553 - 563。doi: 10.1016 / j.phrs.2010.01.012
杜邦、E。,Canu, M. H., Stevens, L., and Falempin, M. (2005). Effects of a 14-day period of hindpaw sensory restriction on mRNA and protein levels of NGF and BDNF in the hindpaw primary somatosensory cortex.大脑摩尔>,Res。133年,78 - 86。doi: 10.1016 / j.molbrainres.2004.09.019
恩斯特,。,一个lkass, K., Bernard, S., Salehpour, M., Perl, S., Tisdale, J., et al. (2014). Neurogenesis in the striatum of the adult human brain.细胞156年,1072 - 1083。doi: 10.1016 / j.cell.2014.01.044
Farioli-Vecchioli, S。Mattera,。Micheli, L。,Ceccarelli, M., Leonardi, L., Saraulli, D., et al. (2014). Running rescues defective adult neurogenesis by shortening the length of the cell cycle of neural stem and progenitor cells.干细胞32岁,1968 - 1982。doi: 10.1002 / stem.1679
弗斯,J。,Stubbs, B., Vancampfort, D., Schuch, F., Lagopoulos, J., Rosenbaum, S., et al. (2018). Effect of aerobic exercise on hippocampal volume in humans: a systematic review and meta-analysis.科学杂志166年,230 - 238。doi: 10.1016 / j.neuroimage.2017.11.007
Folmes, c, D。,Dzeja, P. P., Nelson, T. J., and Terzic, A. (2012). Metabolic plasticity in stem cell homeostasis and differentiation.细胞干细胞11日,596 - 606。doi: 10.1016 / j.stem.2012.10.002
Gaignier F。,Schenten, V., De Carvalho Bittencourt, M., Gauquelin-Koch, G., Frippiat, J. P., and Legrand-Frossi, C. (2014). Three weeks of murine hindlimb unloading induces shifts from B to T and from th to tc splenic lymphocytes in absence of stress and differentially reduces cell-specific mitogenic responses.《公共科学图书馆•综合》9:e92664。doi: 10.1371 / journal.pone.0092664
Giudice, J。,和Taylor, J. M. (2017). Muscle as a paracrine and endocrine organ.咕咕叫。当今。杂志。34岁49-55。doi: 10.1016 / j.coph.2017.05.005
Givogri, m . I。Bottai D。、朱、h·L。法,S。,Lamorte, G., Brambilla, R., et al. (2008). Multipotential neural precursors transplanted into the metachromatic leukodystrophy brain fail to generate oligodendrocytes but contribute to limit brain dysfunction.Dev。>。340 - 357年。doi: 10.1159 / 000150127
Gritti,。,Bonfanti, L., Doetsch, F., Caille, I., Alvarez-Buylla, A., Lim, D. A., et al. (2002). Multipotent neural stem cells reside into the rostral extension and olfactory bulb of adult rodents.j . >。22日,437 - 445。doi: 10.1523 / jneurosci.22 - 02 - 00437.2002
Gubert F。,Zaverucha-do-Valle, C., Pimentel-Coelho, P. M., Mendez-Otero, R., and Santiago, M. F. (2009). Radial glia-like cells persist in the adult rat brain.大脑Res。1258年,43-52。doi: 10.1016 / j.brainres.2008.12.021
Guure, c, B。,Ibrahim, N. A., Adam, M. B., and Said, S. M. (2017). Impact of physical activity on cognitive decline, dementia, and its subtypes: meta-analysis of prospective studies.生物医学。Int >,2017:9016924。doi: 10.1155 / 2017/9016924
哈洛b P。、Bikle D D。锥,c . M。,和Morey-Holton, E. (1988). Glucocorticoids and inhibition of bone formation induced by skeletal unloading.点。j .杂志。255(6分),E875-E879。doi: 10.1152 / ajpendo.1988.255.6.E875
汉森,a . M。,Harrison, B. C., Young, M. H., Stodieck, L. S., and Ferguson, V. L. (2013). Longitudinal characterization of functional, morphologic, and biochemical adaptations in mouse skeletal muscle with hindlimb suspension.肌肉神经48岁,393 - 402。doi: 10.1002 / mus.23753
桥本,M。荒木,Y。,Takashima, Y., Nogami, K., Uchino, A., Yuzuriha, T., et al. (2017). Hippocampal atrophy and memory dysfunction associated with physical inactivity in community-dwelling elderly subjects: the Sefuri study.大脑Behav。7:e00620。doi: 10.1002 / brb3.620
贾,G。,Fu, Y., and He, C. (2013). Reversible RNA adenosine methylation in biological regulation.趋势麝猫。29日,108 - 115。doi: 10.1016 / j.tig.2012.11.003
Juszczak, g R。,和Stankiewicz, A. M. (2018). Glucocorticoids, genes and brain function.掠夺。Neuropsychopharmacol。医学杂志。精神病学82年,136 - 168。doi: 10.1016 / j.pnpbp.2017.11.020
Kakurin, l . I。,Cherepakhin, M. A., Ushakov, A. S., and Senkevich, Y. A. (1972). Functional insufficiency of the neuromuscular system caused by weightlessness and hypokinesia.生命科学。空间Res。10日,61 - 64。
【年代。,Ohmori, S., Kanda, K., Ito, T., Murata, Y., and Seo, H. (1994). Adrenocortical response to tail-suspension in young and old rats.环绕。地中海。38岁的7 - 12。
Kollmann, K。海勒,G。,Schneckenleithner, C., Warsch, W., Scheicher, R., Ott, R. G., et al. (2013). A kinase-independent function of CDK6 links the cell cycle to tumor angiogenesis.癌症细胞24岁,167 - 181。doi: 10.1016 / j.ccr.2013.07.012
库恩,h·G。,Eisch, A. J., Spalding, K., and Peterson, D. A. (2016). Detection and phenotypic characterization of adult neurogenesis.冷泉哈布。教谕。医学杂志。8:a025981。doi: 10.1101 / cshperspect.a025981
Laaksonen d E。、Lakka h . M。,Salonen, J. T., Niskanen, L. K., Rauramaa, R., and Lakka, T. A. (2002). Low levels of leisure-time physical activity and cardiorespiratory fitness predict development of the metabolic syndrome.糖尿病护理25日,1612 - 1618。doi: 10.2337 / diacare.25.9.1612
Langlet C。,Bastide, B., and Canu, M. H. (2012). Hindlimb unloading affects cortical motor maps and decreases corticospinal excitability.经验,神经。237年,211 - 217。doi: 10.1016 / j.expneurol.2012.06.018
李,j . C。,Yau, S. Y., Lee, T. M. C., Lau, B. W., and So, K. F. (2016). Voluntary wheel running reverses the decrease in subventricular zone neurogenesis caused by corticosterone.细胞移植。25日,1979 - 1986。doi: 10.3727 / 096368916 x692195
刘,H。,Wang, Z. C., Yue, Y., Yu, J. W., Cai, Y., Bai, Y. G., et al. (2014). Simulated microgravity induces an inflammatory response in the common carotid artery of rats.可以。j .杂志。杂志。92年,661 - 668。doi: 10.1139 / cjpp - 2014 - 0066
刘,W。,和Crews, F. T. (2017). Persistent decreases in adult subventricular and hippocampal neurogenesis following adolescent intermittent ethanol exposure.前面。Behav。>。11:151。doi: 10.3389 / fnbeh.2017.00151
卢卡斯,J·J。,Domenico, J., and Gelfand, E. W. (2004). Cyclin-dependent kinase 6 inhibits proliferation of human mammary epithelial cells.摩尔。癌症Res。2,105 - 114。
马费伊,M。,Longa, E., Qaisar, R., Agoni, V., Desaphy, J. F., Camerino, D. C., et al. (2014). Actin sliding velocity on pure myosin isoforms from hindlimb unloaded mice.学报杂志。212年,316 - 329。doi: 10.1111 / apha.12320
Masrour, F F。Peeri, M。,一个zarbayjani, M. A., and Hosseini, M. J. (2018). Voluntary exercise during adolescence mitigated negative the effects of maternal separation stress on the depressive-like behaviors of adult male rats: role of NMDA receptors.Neurochem。Res。doi: 10.1007 / s11064 - 018 - 2519 - 6
Mastrorilli, V。花粉刷,C。,Saraulli, D., Costanzi, M., Scardigli, R., Rouault, J. P., et al. (2017). Physical exercise rescues defective neural stem cells and neurogenesis in the adult subventricular zone of Btg1 knockout mice.大脑结构。功能。222年,2855 - 2876。doi: 10.1007 / s00429 - 017 - 1376 - 4
麦地那,j . M。,和Tabernero, A. (2005). Lactate utilization by brain cells and its role in CNS development.j . >。Res。79年,2 - 10。doi: 10.1002 / jnr.20336
Mohyeldin,。,Garzón-Muvdi, T., and Quiñones-Hinojosa, A. (2010). Oxygen in stem cell biology: a critical component of the stem cell niche.细胞干细胞7,150 - 161。doi: 10.1016 / j.stem.2010.07.007
莫雷,e R。,Sabelman, E. E., Turner, R. T., and Baylink, D. J. (1979). A new rat model simulating some aspects of space flight.生理学家22日,S23-S24。
Morey-Holton, e R。,和Globus, R. K. (2002). Hindlimb unloading rodent model: technical aspects.j:。杂志。92年,1367 - 1377。doi: 10.1152 / japplphysiol.00969.2001
Mysoet, J。,Canu, M. H., Cieniewski-Bernard, C., Bastide, B., and Dupont, E. (2014). Hypoactivity affects IGF-1 level and PI3K/AKT signaling pathway in cerebral structures implied in motor control.《公共科学图书馆•综合》9:e107631。doi: 10.1371 / journal.pone.0107631
细语,o . C。,Schultz, S. A., Oh, J. M., Larson, J., Edwards, D., Cook, D., et al. (2014). Physical activity attenuates age-related biomarker alterations in preclinical AD.神经学83年,1753 - 1760。doi: 10.1212 / WNL.0000000000000964
Pourteymour, S。的时候,K。Holen, T。,Langleite, T., Lee, S., Jensen, J., et al. (2017). Global mRNA sequencing of human skeletal muscle: search for novel exercise-regulated myokines.摩尔。金属底座。6,352 - 365。doi: 10.1016 / j.molmet.2017.01.007
Rogatzki, m . J。,Ferguson, B. S., Goodwin, M. L., and Gladden, L. B. (2015). Lactate is always the end product of glycolysis.前面。>。九22。doi: 10.3389 / fnins.2015.00022
Safdar,。,和Tarnopolsky, M. A. (2018). Exosomes as mediators of the systemic adaptations to endurance exercise.冷泉哈布。教谕。地中海。8:a029827。doi: 10.1101 / cshperspect.a029827
Sanai, N。,Nguyen, T., Ihrie, R. A., Mirzadeh, Z., Tsai, H. H., Wong, M., et al. (2011). Corridors of migrating neurons in the human brain and their decline during infancy.自然478年,382 - 386。doi: 10.1038 / nature10487
Sanai, N。,Tramontin, A. D., Quiñones-Hinojosa, A., Barbaro, N. M., Gupta, N., Kunwar, S., et al. (2004). Unique astrocyte ribbon in adult human brain contains neural stem cells but lacks chain migration.自然427年,740 - 744。doi: 10.1038 / nature02301
Santilli G。,Lamorte, G., Carlessi, L., Ferrari, D., Rota Nodari, L., Binda, E., et al. (2010). Mild hypoxia enhances proliferation and multipotency of human neural stem cells.《公共科学图书馆•综合》5:e8575。doi: 10.1371 / journal.pone.0008575
尊敬。,和Hochedlinger, K. (2013). The sox family of transcription factors: versatile regulators of stem and progenitor cell fate.细胞干细胞12、15 - 30。doi: 10.1016 / j.stem.2012.12.007
沈问。,Wang, Y., Kokovay, E., Lin, G., Chuang, S. M., Goderie, S. K., et al. (2008). Adult SVZ stem cells lie in a vascular niche: a quantitative analysis of niche cell-cell interactions.细胞干细胞3,289 - 300。doi: 10.1016 / j.stem.2008.07.026
斯奈德,j·S。,Cahill, S. P., and Frankland, P. W. (2017). Running promotes spatial bias independently of adult neurogenesis.海马体27日,871 - 882。doi: 10.1002 / hipo.22737
、L。,Csete, M., Lee, S. H., Kabbani, N., Walikonis, J., Wold, B., et al. (2000). Enhanced proliferation, survival, and dopaminergic differentiation of CNS precursors in lowered oxygen.j . >。20岁,7377 - 7383。doi: 10.1523 / jneurosci.20 07377.2000 - 19
太阳,x Q。,Xu, Z. P., Zhang, S., Cao, X. S., and Liu, T. S. (2009). Simulated weightlessness aggravates hypergravity-induced impairment of learning and memory and neuronal apoptosis in rats.Behav。大脑Res。199年,197 - 202。doi: 10.1016 / j.bbr.2008.11.035
泰勒,S。,Wakem, M., Dijkman, G., Alsarraj, M., and Nguyen, M. (2010). A practical approach to RT-qPCR-Publishing data that conform to the MIQE guidelines.方法S1-S5。doi: 10.1016 / j.ymeth.2010.01.005
Tigan, a S。,Bellutti, F., Kollmann, K., Tebb, G., and Sexl, V. (2016). CDK6-a review of the past and a glimpse into the future: from cell-cycle control to transcriptional regulation.致癌基因35岁,3083 - 3091。doi: 10.1038 / onc.2015.407
van den河岸,s。Middeldorp, J。张,c, E。,Curtis, M. A., Leonard, B. W., Mastroeni, D., et al. (2010). Longterm quiescent cells in the aged human subventricular neurogenic system specifically express GFAP-delta.衰老细胞9日,313 - 326。doi: 10.1111 / j.1474-9726.2010.00556.x
Vandesompele, J。现成的,K。Pattyn F。波,B。,Van Roy, N., De Paepe, A., et al. (2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes.基因组医学杂志3:RESEARCH0034。doi: 10.1186 / gb - 2002 - 3 - 7 - research0034
van Praag, H。,Kempermann, G., and Gage, F. H. (1999). Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus.Nat。>。2,266 - 270。doi: 10.1038/6368
van Praag, H。Shubert, T。赵,C。,和Gage, F. H. (2005). Exercise enhances learning and hippocampal neurogenesis in aged mice.j . >。25日,8680 - 8685。doi: 10.1523 / jneurosci.1731 - 05.2005
王,H。,Wei, L., Li, C., Zhou, J., and Li, Z. (2015). CDK5RAP1 deficiency induces cell cycle arrest and apoptosis in human breast cancer cell line by the ROS/JNK signaling pathway.肿瘤防治杂志。代表。33岁,1089 - 1096。doi: 10.3892 / or.2015.3736
王,T。,Chen, H., Lv, K., Ji, G., Zhang, Y., Wang, Y., et al. (2017). iTRAQ-based proteomics analysis of hippocampus in spatial memory deficiency rats induced by simulated microgravity.j .蛋白质组学160年,64 - 73。doi: 10.1016 / j.jprot.2017.03.013
魏,f . Y。周,B。,Suzuki, T., Miyata, K., Ujihara, Y., Horiguchi, H., et al. (2015). Cdk5rap1-mediated 2-methylthio modification of mitochondrial tRNAs governs protein translation and contributes to myopathy in mice and humans.细胞金属底座。21日,428 - 442。doi: 10.1016 / j.cmet.2015.01.019
Whitham, M。,Parker, B. L., Friedrichsen, M., Hingst, J. R., Hjorth, M., Hughes, W. E., et al. (2018). Extracellular vesicles provide a means for tissue crosstalk during exercise.细胞金属底座。27日,237 - 251 e234。doi: 10.1016 / j.cmet.2017.12.001
Yasuhara, T。Hara, K。真希,M。,Matsukawa, N., Fujino, H., Date, I., et al. (2007). Lack of exercise, via hindlimb suspension, impedes endogenous neurogenesis.神经科学149年,182 - 191。doi: 10.1016 / j.neuroscience.2007.07.045
关键词:神经干细胞、运动不活跃,神经退行性疾病,宇航员,后肢卸货,神经发生,Cdk5RAP1,新陈代谢
引用:阿达米R, Pagano J,科伦坡,Platonova N, Recchia D, Chiaramonte R, Bottinelli R, Canepari M和Bottai D(2018)减少运动神经系统疾病:对神经干细胞特性的影响。前面。>。12:336。doi: 10.3389 / fnins.2018.00336
收到:2018年2月02;接受:2018年4月30日;
发表:2018年5月23日。
编辑:
文森特Tropepe加拿大多伦多大学版权阿达米©2018,Pagano,科伦坡,Platonova Recchia) Chiaramonte, Bottinelli, Canepari Bottai。这是一个开放分布式根据文章知识共享归属许可(CC)。使用、分发或复制在其他论坛是允许的,提供了原始作者(年代)和版权所有者认为,最初发表在这个期刊引用,按照公认的学术实践。没有使用、分发或复制是不符合这些条件的允许。
*通信:Daniele Bottai,daniele.bottai@unimi.it
__现在地址:杰西卡·Pagano医学生物技术和转化医学、意大利米兰