

CD8 + T细胞


T细胞淋巴细胞构成细胞过程中发挥重要作用适应性免疫。当T细胞受体(tcr)上发现幼稚T细胞识别其特定的抗原与主要组织相容性复合体(MHC)类蛋白质,他们成熟和扩散到专门的T细胞类型的子集执行免疫功能的关键。在本文中,我们关注CD8 + T细胞通过描述他们的免疫表型和细胞介导的免疫功能背后的过程。
CD8 + T细胞是什么?
通常被称为细胞毒性T淋巴细胞(ctl), CD8 + T细胞表达CD8属于一个群面。CD8是二聚的受体,使CD8 + T细胞识别我蛋白质肽MHC类提供的。主要组织相容性复合体(MHC)是一个包含许多地区的DNA基因编码糖蛋白分子免疫系统识别的关键的外国材料。
有两个主要类别的MHC分子。他们之间存在重要差异。
在胸腺,成功绑定一个天真的CD8 + T细胞和一个之间抗原呈递细胞(APC)刺激不成熟T细胞成为激活CD8 + T细胞与细胞毒性功能。与细胞毒性细胞内病原体的杀戮和功能肿瘤细胞ctl属于一组专门的淋巴细胞参与细胞介导获得性免疫。1虽然ctl免疫系统功能是必不可少的,ctl的失调可以导致瀑特异性自身免疫性疾病的发病机制2像糖尿病和关节炎。
CD8 + T细胞从何而来?
所有的T细胞来源于淋巴祖细胞从骨髓迁移到胸腺。胸腺房子不成熟T细胞尚未开发所需抗原T细胞受体识别。在感染期间,天真的CD8 + T细胞最初刺激淋巴器官内与装甲运兵车交互。CD8 + T细胞的发展
一旦不成熟的T细胞迁移到胸腺,他们准备发展成为CD8 +细胞。祖T细胞淋巴细胞第一次经历许多DNA重组,产生细胞的多样性从外国或胞内病原体识别抗原的能力。识别多样性是依赖于随机DNA重组TCR基因的数量导致独特的表达,抗原受体。一旦确定细胞像CD8功能稳定,不成熟的T细胞已经准备好接受一个antigen-mediated过程被称为克隆选择。积极的选择开始当细胞受体结合与MHC抗原在复杂类蛋白质由胸腺细胞。如果这个绑定事件成功CD8、新激活的CD8 + T细胞将分化成成熟的ctl。不成熟的T细胞不能充分结合antigen-MHC类我复杂的在这个阶段会进行细胞凋亡。负选择发生在不成熟的T细胞结合太强烈,自体抗原。消极的选择以T细胞程序性凋亡,这消除了他们成为autoreactive攻击健康细胞的能力。
CD8 + T细胞激活
天真的CD8 + T细胞,进行积极的选择需要额外的信令流程来确定其长期的命运。3例如,CD4 +辅助T细胞可以调节装甲运兵车呈现较强的抗原信号天真的CD8 + T细胞。辅助T细胞和树突细胞(dc)还可以通过costimulatory调解细胞毒性T细胞激活信号如CD80/86以及细胞因子il - 12。同时激活的tcr costimulatory T细胞代数余子式许可激活CD8 + T细胞增殖为一个扩展的人口,其中包含效应和记忆T细胞类型。这种扩张的积极选择的CD8 + T细胞的高度特异性ctl是克隆选择的最后一步。细胞毒性T细胞做什么?
一旦CD8 + T细胞被激活,他们接受级联的复制和分化的步骤质数他们有针对性的免疫反应。ctl物理接触目标感染或恶性肿瘤细胞诱导细胞凋亡,细胞程序性死亡的一种形式,由细胞毒性酶和分子。
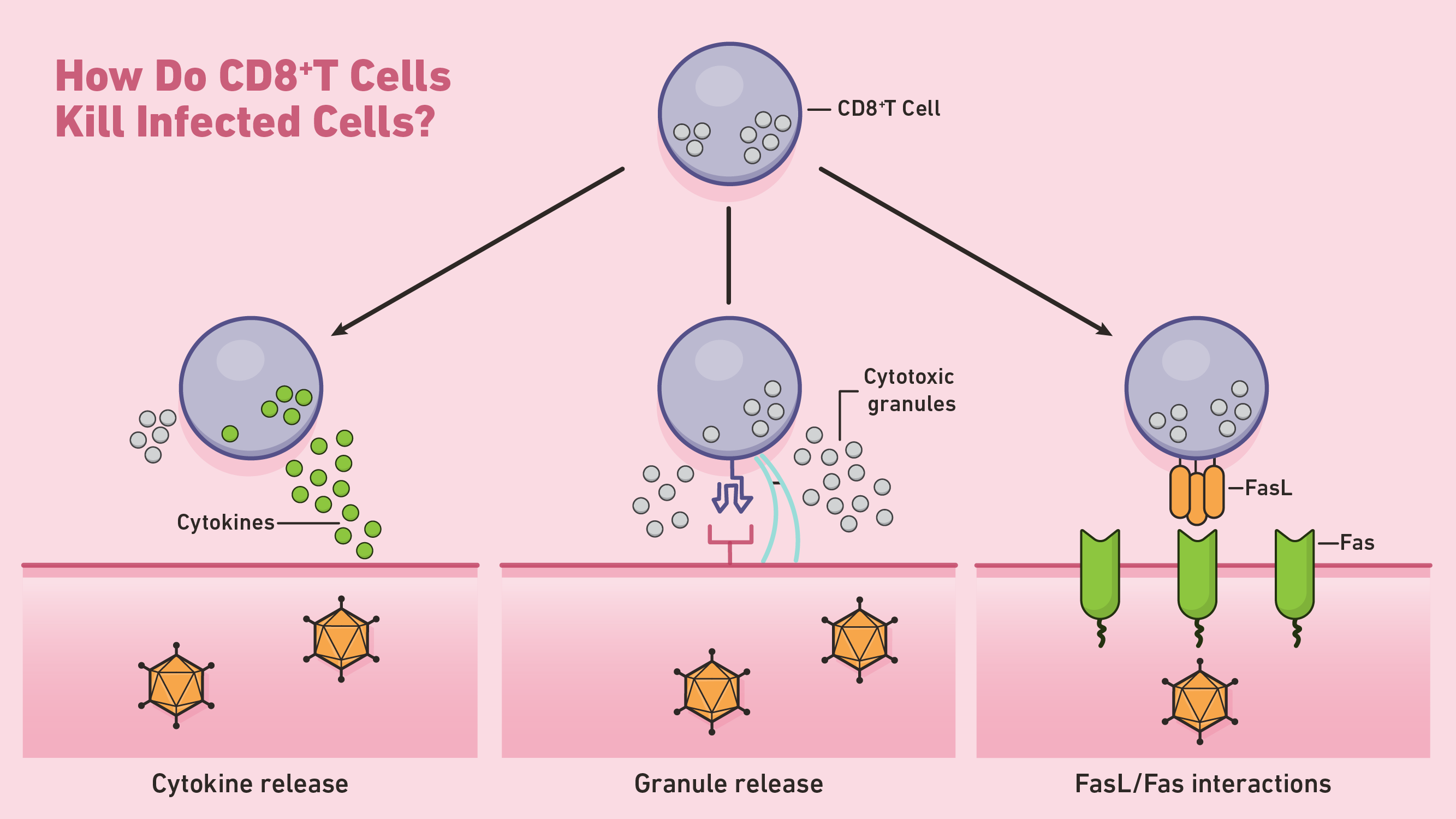
图1:CD8 + T细胞杀死感染细胞如何?
一种CD8 + T细胞启动细胞凋亡是通过穿孔素的分泌和granzymes,两种类型的细胞毒性蛋白。穿孔素细胞溶解的蛋白质形成毛孔在目标细胞的细胞膜。ctl使用这些毛孔直接granzymes的释放,一种丝氨酸蛋白酶,继续胞液中细胞凋亡的过程。这些蛋白酶关闭目标细胞,病毒和细胞的蛋白质参与正常细胞的维护。有针对性的细胞发生凋亡然后由附近的吞噬细胞清除。
ctl通过Fas-Fas配体的相互作用也可以诱导细胞凋亡。每当Fas配体(FasL)结合Fas受体靶向细胞,一系列的信号分子通过激活半胱天冬酶启动细胞凋亡蛋白酶。这些还进行大量的蛋白水解作用在细胞凋亡的后期。
除了直接细胞凋亡,CD8 + T细胞可以杀死靶细胞间接通过释放细胞因子等因素TNF -α。例如,1型CD8 + T细胞(Tc1)可以释放IFN -γ,能够抑制病毒复制的细胞因子,提高特异性抗原的表达。4结束时,他们的主要反应,激活CD8 + T细胞通常死的细胞凋亡。
CD8 + T细胞亚型
天真的CD8 + T细胞分化成效应或记忆细胞类型执行特定角色,调节适应性免疫反应。T细胞诱导成为效应细胞分化成Tc1、Tc2细胞。Tc1细胞获得的能力杀死目标细胞直接或间接一旦他们成为刺激等细胞因子和白介素- 2。此外,细胞毒性Tc1细胞可以释放干扰素- γ 或肿瘤坏死因子- α 进一步调节宿主免疫反应。Tc2 T细胞调节效应和辅助免疫反应通过分泌细胞因子il - 4、il - 10和IL-13 IL-5。虽然Tc2细胞具备执行细胞毒性功能,这个子集的T细胞通常是不那么有效Tc1同行在感染。除了这些良好的亚型,新的亚型,如Tc9 Tc17和Tc22近年来已确定,每个和产生不同的细胞因子引起的(图2)。
图2:CD8 + T细胞亚型包括Tc1、Tc2, Tc9, Tc17 Tc22。
内存CD8 + T细胞通常是确认后的收缩效应细胞群。某些效应细胞保持收缩后,成为记忆细胞。内存子集的CD8 + T细胞可以分为三个不同的组:中央记忆细胞,效应记忆细胞和tissue-resident记忆细胞。他们被认为是主要来源于Tc1子集1。这些子集的特点是其功能的差异,增殖能力和位置。中央记忆细胞(中医)lymphoid-residing淋巴细胞,通常通过快速扩散对继发感染。在人类中,中医细胞表型特征的co-expression表面标记CD127 CD27和CD28。效应记忆细胞(Tem)流通全身准备寻找并杀死antigen-matching靶细胞。与中医同行不同的是,这些效应记忆细胞没有表现出一个健壮的增殖和自我更新能力,以应对感染。最后,tissue-resident记忆细胞(Trm)成熟ctl“备用”继发感染损伤或原始网站的入侵。
vs CD4 + CD8 + T细胞
CD4 +淋巴细胞是一群“助手”T细胞调节其他免疫细胞活动通过直接和间接的机制。天真的CD4 + T细胞位于胸腺,直到他们被装甲运兵车刺激表达MHC II级目标抗原结合蛋白。这种蛋白质复杂的成功绑定激活CD4 + T细胞开始分化成一个辅助细胞类型。尽管CD4 + T细胞激活不执行细胞毒性功能像CD8 + T细胞,成熟后为辅助T细胞可以调解体液或细胞介导的免疫反应通过细胞因子的分泌。5比较 |
CD8 + T细胞 |
CD4 + T细胞 |
类型 |
细胞毒性或“杀手”细胞 |
“助手”细胞 |
关键功能(年代) |
摧毁受感染或恶性肿瘤细胞 |
调节免疫细胞的活动 |
TCR-MHC绑定复杂 |
CD8 +类MHC I |
CD4 + MHC II级 |
数量的子类型 |
五 |
六个 |
CD8 + T细胞在哪里适合更广泛的免疫反应吗?
CD8 + t细胞调节取决于刺激抑制机制由tcr的集合。当CD8 + T细胞呈现特异性抗原,他们经历许多克隆扩张,产生分化细胞类型所需的一个适应性免疫应答。独立通过识别抗原的演讲,CD8 + T细胞可以间接攻击病原体或肿瘤的细胞通过细胞因子的分泌。6然而,不受监管的CTL活动可以有助于对健康细胞的自体免疫反应。CD8 + t细胞过程的更大理解在这种情况下,可以提高我们的能力,设计有效的疫苗或实时操纵细胞介导的免疫反应。
引用
1。张,N,贝文,乔丹。CD8 + T细胞:步兵的免疫系统。免疫力。2011年,35 (2):161 - 168。doi:10.1016 / j.immuni.2011.07.010
2。吴罗邓Q, Y, Chang C, H,丁Y和小r .新兴后生CD8 + T细胞在自身免疫性疾病中的作用:一项系统回顾。前面。Immunol。2019;10:856。doi:10.3389 / fimmu.2019.00856
3所示。霍耶年代,Prommersberger年代,Pfeiffer IA Schuler-Thurner B,舒勒克,Dorrie J, Schaft n .并发交互DCs的CD4(+)和CD8 (+) T细胞改善二级CTL扩张:需要三个探戈。欧元。j . Immunol。2014年,44 (12):3543 - 59。doi:10.1002 / eji.201444477
4所示。τG,罗斯曼IFN-γ受体的生物学功能。过敏。1999年,54 (12):1233 - 1251。doi:10.1034 / j.1398-9995.1999.00099.x
5。我们朱J,保罗。(2008)。CD4 T细胞:命运、函数和缺点。血。2008,112 (5):1557 - 69。doi:10.1182 / - 2008 - 05 - 078154血
6。Berg再保险,福尔曼j . CD8 T细胞的作用在先天免疫和抗原非特异性保护。当今Immunol。2006;18 (3):338 - 343。doi:10.1016 / j.coi.2006.03.010

